AG Johansson
Forschungsgebiete
Licht kontrolliert als externes Signal viele Aspekte des Wachstums und der Entwicklung im Verlauf des pflanzlichen Lebenszyklus, einschließlich Keimung, De-Etiolierung, Schattenflucht, Bewegung von Chloroplasten, Phototropismus, Einschwingen der cirkadianen Uhr, und Blühzeitpunkt. Die De-Etiolierung von Keimlingen ist eine der stärksten Antworten auf einen einzelnen Stimulus in der Natur, und daher ist es nicht verwunderlich, dass diese Antwort einen großen Anteil des pflanzlichen Transkriptoms betrifft. Dies wird hauptsächlich durch die Rot- und Blaulichtrezeptoren vermittelt (Phytochrome und Cryptochrome). Diese Photorezeptoren regulieren die Transkription jedoch nicht direkt, sondern über ein komplexes Netzwerk von positiven und negativen Regulatoren der Photomorphogenese.
Vor mehr als zwei Jahrzehnten wurde COP1 aufgrund seines photomorphogenetischen Phänotyps in der Dunkelheit als wichtiger negativer Regulator der Lichtsignaltransduktion identifiziert. Im Dunkeln wirkt COP1 als E3-Ubiquitinligase, die mehrere positive Faktoren der Photomorphogenese, insbesondere den bZIP-Transkriptionsfaktor HY5, zum Abbau markiert. Bei Belichtung wird die Aktivität von COP1 durch die Photorezeptoren gehemmt und das Protein aus dem Zellkern ausgeschlossen, was in der Anreicherung von HY5 und in der Folge zur De-Etiolierung führt.
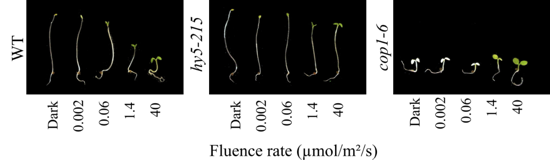
Fotos von Keimlingen, die 7 Tage lang im Dunkeln oder in zunehmender Dosis von Rotlicht angezogen wurden. Zunehmende Lichtintensitäten führen beim Wildtyp zur Öffnung und Entfaltung der Keimblätter, Hemmung des Längenwachstums des Hypokotyls und Chlorophyllanreicherung. Die hy5-Mutante ist unempfindlich gegenüber Licht, während die cop1-Mutante bereits in der Dunkelheit photomorphogenetisch ist.
In der AG Johansson interessieren wir uns für die molekularen Ereignisse, die zwischen der Perzeption von Licht durch die Photorezeptoren und der darauf folgenden Transkriptionsantwort während der De-Etiolierung stattfinden. Wir arbeiten mit einer Gruppe von B-Box-Zinkfingerproteinen, die physisch mit COP1 und / oder HY5 interagieren. Wie durch diese Interaktionen nahegelegt, modulieren diese B-Box-Proteine tatsächlich den Lichtsignalweg, was sich in Mutanten durch geänderte Hemmung des Längenwachstums des Hypokotyls und die Anreicherung von Anthocyan zeigt. Im Moment untersuchen wir die Ähnlichkeiten und Unterschiede zwischen diesen B-Box-Proteinen und versuchen den Mechanismus herauszufinden, mittels dem sie die Aktivität von HY5 modulieren.


